
Photoheterotrophs (Gk: photo = light, hetero = (an)other, troph = nourishment) are heterotrophic phototrophs – that is, they are organisms that use light for energy, but cannot use carbon dioxide as their sole carbon source. Consequently, they use organic compounds from the environment to satisfy their carbon requirements; these compounds include carbohydrates, fatty acids, and alcohols. Examples of photoheterotrophic organisms include purple non-sulfur bacteria, green non-sulfur bacteria, and heliobacteria. These microorganisms are ubiquitous in aquatic habitats, occupy unique niche-spaces, and contribute to global biogeochemical cycling. Recent research has also indicated that the oriental hornet and some aphids may be able to use light to supplement their energy supply.
Research
Studies have shown that mammalian mitochondria can also capture light and synthesize ATP when mixed with pheophorbide, a light-capturing metabolite of chlorophyll. Research demonstrated that the same metabolite when fed to the worm Caenorhabditis elegans leads to increase in ATP synthesis upon light exposure, along with an increase in life span.
Furthermore, inoculation experiments suggest that mixotrophic Ochromonas danica (i.e., Golden algae)—and comparable eukaryotes—favor photoheterotrophy in oligotrophic (i.e., nutrient-limited) aquatic habitats. This preference may increase energy-use efficiency and growth by reducing investment in inorganic carbon fixation (e.g., production of autotrophic machineries such as RuBisCo and PSII).
Metabolism
Photoheterotrophs generate ATP using light, in one of two ways: they use a bacteriochlorophyll-based reaction center, or they use a bacteriorhodopsin. The chlorophyll-based mechanism is similar to that used in photosynthesis, where light excites the molecules in a reaction center and causes a flow of electrons through an electron transport chain (ETS). This flow of electrons through the proteins causes hydrogen ions to be pumped across a membrane. The energy stored in this proton gradient is used to drive ATP synthesis. Unlike in photoautotrophs, the electrons flow only in a cyclic pathway: electrons released from the reaction center flow through the ETS and return to the reaction center. They are not utilized to reduce any organic compounds. Purple non-sulfur bacteria, green non-sulfur bacteria, and heliobacteria are examples of bacteria that carry out this scheme of photoheterotrophy.
Other organisms, including halobacteria and flavobacteria and vibrios have purple-rhodopsin-based proton pumps that supplement their energy supply. The archaeal version is called bacteriorhodopsin, while the eubacterial version is called proteorhodopsin. The pump consists of a single protein bound to a Vitamin A derivative, retinal. The pump may have accessory pigments (e.g., carotenoids) associated with the protein. When light is absorbed by the retinal molecule, the molecule isomerises. This drives the protein to change shape and pump a proton across the membrane. The hydrogen ion gradient can then be used to generate ATP, transport solutes across the membrane, or drive a flagellar motor. One particular flavobacterium cannot reduce carbon dioxide using light, but uses the energy from its rhodopsin system to fix carbon dioxide through anaplerotic fixation. The flavobacterium is still a heterotroph as it needs reduced carbon compounds to live and cannot subsist on only light and CO2. It cannot carry out reactions in the form of
where H2D may be water, H2S or another compound/compounds providing the reducing electrons and protons; the 2D + H2O pair represents an oxidized form.
However, it can fix carbon in reactions like:
where malate or other useful molecules are otherwise obtained by breaking down other compounds by
- carbohydrate + O2 → malate + CO2 + energy.
This method of carbon fixation is useful when reduced carbon compounds are scarce and cannot be wasted as CO2 during interconversions, but energy is plentiful in the form of sunlight.
Ecology

Distribution and Niche Partitioning
Photoheterotrophs—either 1) cyanobacteria (i.e. facultative heterotrophs in nutrient-limited environments like Synechococcus and Prochlorococcus), 2) aerobic anoxygenic photoheterotrophic bacteria (AAP; employing bacteriochlorophyll-based reaction centers), 3) proteorhodopsin (PR)-containing bacteria and archaea, and 4) heliobacteria (i.e., the only phototroph with bacteriochlorophyll g pigments, or Gram-positive membrane) are found in various aquatic habitats including oceans, stratified lakes, rice fields, and environmental extremes.
In surface "oceans", up to 10% of bacterial cells are capable of AAP, whereas greater than 50% of net marine microorganisms house PR—reaching up to 90% in coastal biomes. As demonstrated in inoculation experiments, photoheterotrophy may provide these planktonic microbes competitive advantages 1) relative to chemoheterotrophs in oligotrophic (i.e., nutrient-poor) environments via increased nutrient use-efficiency (i.e., organic carbon fuels biosynthesis, excessively, versus energy production) and 2) by eliminating investment in physiologically costly autotrophic enzymes/complexes (RuBisCo and PSII). Furthermore, in Arctic oceans, AAP and PR photoheterotrophs are prominent in ice-covered regions during wintertime per light scarcity. Lastly, seasonal turnover has been observed in marine AAPs as ecotypes (i.e., genetically similar taxa with differing functional trait and/or environmental preferences) segregate into temporal niches.
In stratified (i.e., euxinic) lakes, photoheterotrophs—alongside other anoxygenic phototrophs (e.g., purple/green sulfur bacteria fixing carbon dioxide via electron donors such as ferrous iron, sulfide, and hydrogen gas)—often occupy the chemocline in the water column and/or sediments. In this zone, dissolved oxygen is reduced, light is limited to long wavelengths (e.g., red and infrared) left-over by oxygenic phototrophs (e.g., cyanobacteria), and anaerobic metabolisms (i.e., those occurring in the absence of oxygen) begin introducing sulfide and bioavailable nutrients (e.g., organic carbon, phosphate, and ammonia) through upward diffusion.
Heliobacteria are obligate anaerobes primarily located in rice fields, where low sulfide concentrations prevent competitive exclusion of purple/green sulfur bacteria. These waterlogged environments may facilitate symbiotic relationships between heliobacteria and rice plants as fixed nitrogen—from the former—is exchanged for carbon-rich root exudates.
Observation studies have characterized photoheterotrophs (e.g., Green non-sulfur bacteria such as Chloroflexi and AAPs) within photosynthetic mats at environmental extremes (e.g., hot springs and hypersaline lagoons). Notably, temperature and pH drive anoxygenic phototroph community composition in Yellowstone National Park's geothermal features. In addition, various, light-dependent niches in the Great Salk Lake's hypersaline mats support phototrophic diversity as microbes optimize energy production and combat osmotic stress.
Biogeochemical Cycling
Photoheterotrophs influence global carbon cycling by assimilating dissolved organic carbon (DOC). Therefore, when harvesting light-energy, carbon is maintained in the microbial loop without corresponding respiration (i.e., carbon dioxide release to the atmosphere as DOC is oxidized to fuel energy production). This disconnect, the discovery of facultative photoheterotrophs (e.g., AAPs with flexible energy sources), and previous measurements taken in the dark (i.e., to avoid skewed oxygen consumption values due to photooxidation, UV light, and oxygenic photosynthesis) lead to overestimated aquatic CO2 emissions. For example, a 15.2% decrease in community respiration was observed in Cep Lake, Czechia—alongside preferential glucose and pyruvate uptake—is attributed to facultative photoheterotrophs preferring light-energy during the daytime, given fitness benefits mentioned previously.
Flowchart
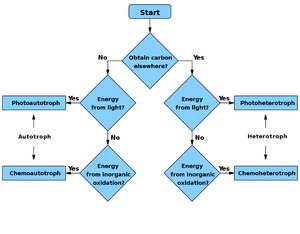
|
Energy source
Carbon source
|
Chemotroph | Phototroph |
|---|---|---|
| Autotroph | Chemoautotroph | Photoautotroph |
| Heterotroph | Chemoheterotroph | Photoheterotroph |
See also
Sources
"Microbiology Online" (textbook). University of Wisconsin, Madison.