




Thermogenin (called uncoupling protein by its discoverers and now known as uncoupling protein 1, or UCP1) is a mitochondrial carrier protein found in brown adipose tissue (BAT). It is used to generate heat by non-shivering thermogenesis, and makes a quantitatively important contribution to countering heat loss in babies which would otherwise occur due to their high surface area-volume ratio.
Mechanism
UCP1 belongs to the UCP family which are transmembrane proteins that decrease the proton gradient generated in oxidative phosphorylation. They do this by increasing the permeability of the inner mitochondrial membrane, allowing protons that have been pumped into the intermembrane space to return to the mitochondrial matrix. UCP1-mediated heat generation in brown fat uncouples the respiratory chain, allowing for fast substrate oxidation with a low rate of ATP production. UCP1 is related to other mitochondrial metabolite transporters such as the adenine nucleotide translocator, a proton channel in the mitochondrial inner membrane that permits the translocation of protons from the mitochondrial intermembrane space to the mitochondrial matrix. UCP1 is restricted to brown adipose tissue, where it provides a mechanism for the enormous heat-generating capacity of the tissue.
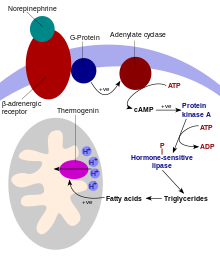
UCP1 is activated in the brown fat cell by fatty acids and inhibited by nucleotides. Fatty acids are released by the following signaling cascade: Sympathetic nervous system terminals release Norepinephrine onto a Beta-3 adrenergic receptor on the plasma membrane. This activates adenylyl cyclase, which catalyses the conversion of ATP to cyclic AMP (cAMP). cAMP activates protein kinase A, causing its active C subunits to be freed from its regulatory R subunits. Active protein kinase A, in turn, phosphorylates triacylglycerol lipase, thereby activating it. The lipase converts triacylglycerols into free fatty acids, which activate UCP1, overriding the inhibition caused by purine nucleotides (GDP and ADP). During the termination of thermogenesis, thermogenin is inactivated and residual fatty acids are disposed of through oxidation, allowing the cell to resume its normal energy-conserving state.

UCP1 is very similar to the ATP/ADP Carrier protein, or Adenine Nucleotide Translocator (ANT). The proposed alternating access model for UCP1 is based on the similar ANT mechanism. The substrate comes in to the half open UCP1 protein from the cytoplasmic side of the membrane, the protein closes the cytoplasmic side so the substrate is enclosed in the protein, and then the matrix side of the protein opens, allowing the substrate to be released into the mitochondrial matrix. The opening and closing of the protein is accomplished by the tightening and loosening of salt bridges at the membrane surface of the protein. Substantiation for this modelling of UCP1 on ANT is found in the many conserved residues between the two proteins that are actively involved in the transportation of substrate across the membrane. Both proteins are integral membrane proteins, localized to the inner mitochondrial membrane, and they have a similar pattern of salt bridges, proline residues, and hydrophobic or aromatic amino acids that can close or open when in the cytoplasmic or matrix state.
Evolution
UCP1 is expressed in brown adipose tissue, which is functionally found only in eutherians. The UCP1, or thermogenin, gene likely arose in an ancestor of modern vertebrates, but did not initially allow for our vertebrate ancestor to use non-shivering thermogenesis for warmth. It wasn't until heat generation was adaptively selected for in placental mammal descendants of this common ancestor that UCP1 evolved its current function in brown adipose tissue to provide additional warmth. While UCP1 plays a key thermogenic role in wide range placental mammals, particularly those with small body size and those that hibernate, the UCP1 gene has lost functionality in several large-bodied lineages (e.g. horses, elephants, sea cows, whales and hyraxes) and lineages with low metabolic rates (e.g. pangolins, armadillos, sloths and anteaters). Recent discoveries of non-heat-generating orthologues of UCP1 in fish and marsupials, other descendants of the ancestor of modern vertebrates, show that this gene was passed on to all modern vertebrates, but aside from placental mammals, none have heat producing capability. This further suggests that UCP1 had a different original purpose and in fact phylogenetic and sequence analyses indicate that UCP1 is likely a mutated form of a dicarboxylate carrier protein that adapted for thermogenesis in placental mammals.
History
Researchers in the 1960s investigating brown adipose tissue, found that in addition to producing more heat than typical of other tissues, brown adipose tissue seemed to short circuit, or uncouple, respiration coupling. Uncoupling protein 1 was discovered in 1976 by David G. Nicholls, Vibeke Bernson, and Gillian Heaton, and the discovery was published in 1978 and shown to be the protein responsible for this uncoupling effect. UCP1 was later purified for the first time in 1980 and was first cloned in 1988.
Uncoupling protein two (UCP2), a homolog of UCP1, was identified in 1997. UCP2 localizes to a wide variety of tissues, and is thought to be involved in regulating reactive oxygen species (ROS). In the past decade, three additional homologs of UCP1 have been identified, including UCP3, UCP4, and UCP5 (also known as BMCP1 or SLC25A14).
Clinical relevance
Methods of delivering UCP1 to cells by gene transfer therapy or methods of its upregulation have been an important line of enquiry in research into the treatment of obesity, due to their ability to dissipate excess metabolic stores.
See also
- 2,4-Dinitrophenol (A synthetic small-molecule proton shuttle with similar effects)
Further reading
- Macher, Gabriel; Koehler, Melanie; Rupprecht, Anne; Kreiter, Jürgen; Hinterdorfer, Peter; Pohl, Elena E. (March 2018). "Inhibition of mitochondrial UCP1 and UCP3 by purine nucleotides and phosphate". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (3): 664–672. doi:10.1016/j.bbamem.2017.12.001. PMC 6118327. PMID 29212043.
- Urbánková, Eva; Voltchenko, Anna; Pohl, Peter; Ježek, Petr; Pohl, Elena E. (29 August 2003). "Transport Kinetics of Uncoupling Proteins". Journal of Biological Chemistry. 278 (35): 32497–32500. doi:10.1074/jbc.M303721200. PMID 12826670.
- Ricquier D, Bouillaud F (Jan 2000). "The uncoupling protein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP". The Biochemical Journal. 345 Pt 2 (2): 161–79. doi:10.1042/0264-6021:3450161. PMC 1220743. PMID 10620491.
- Muzzin P (Apr 2002). "The uncoupling proteins". Annales d'Endocrinologie. 63 (2 Pt 1): 106–10. PMID 11994670.
- Del Mar Gonzalez-Barroso M, Ricquier D, Cassard-Doulcier AM (Oct 2000). "The human uncoupling protein-1 gene (UCP1): present status and perspectives in obesity research". Obesity Reviews. 1 (2): 61–72. doi:10.1046/j.1467-789x.2000.00009.x. PMID 12119988. S2CID 30231289.
- Cassard AM, Bouillaud F, Mattei MG, Hentz E, Raimbault S, Thomas M, Ricquier D (Jul 1990). "Human uncoupling protein gene: structure, comparison with rat gene, and assignment to the long arm of chromosome 4". Journal of Cellular Biochemistry. 43 (3): 255–64. doi:10.1002/jcb.240430306. PMID 2380264. S2CID 31128860.
- Bouillaud F, Villarroya F, Hentz E, Raimbault S, Cassard AM, Ricquier D (Jul 1988). "Detection of brown adipose tissue uncoupling protein mRNA in adult patients by a human genomic probe". Clinical Science. 75 (1): 21–7. doi:10.1042/cs0750021. PMID 3165741.
- Oppert JM, Vohl MC, Chagnon M, Dionne FT, Cassard-Doulcier AM, Ricquier D, Pérusse L, Bouchard C (Aug 1994). "DNA polymorphism in the uncoupling protein (UCP) gene and human body fat". International Journal of Obesity and Related Metabolic Disorders. 18 (8): 526–31. PMID 7951471.
- Clément K, Ruiz J, Cassard-Doulcier AM, Bouillaud F, Ricquier D, Basdevant A, Guy-Grand B, Froguel P (Dec 1996). "Additive effect of A-->G (-3826) variant of the uncoupling protein gene and the Trp64Arg mutation of the beta 3-adrenergic receptor gene on weight gain in morbid obesity". International Journal of Obesity and Related Metabolic Disorders. 20 (12): 1062–6. PMID 8968850.
- Schleiff E, Shore GC, Goping IS (Mar 1997). "Human mitochondrial import receptor, Tom20p. Use of glutathione to reveal specific interactions between Tom20-glutathione S-transferase and mitochondrial precursor proteins". FEBS Letters. 404 (2–3): 314–8. doi:10.1016/S0014-5793(97)00145-2. PMID 9119086. S2CID 29177508.
- Urhammer SA, Fridberg M, Sørensen TI, Echwald SM, Andersen T, Tybjaerg-Hansen A, Clausen JO, Pedersen O (Dec 1997). "Studies of genetic variability of the uncoupling protein 1 gene in Caucasian subjects with juvenile-onset obesity". The Journal of Clinical Endocrinology and Metabolism. 82 (12): 4069–74. doi:10.1210/jcem.82.12.4414. PMID 9398715.
- Jezek P, Urbánková E (Jan 2000). "Specific sequence of motifs of mitochondrial uncoupling proteins". IUBMB Life. 49 (1): 63–70. doi:10.1080/713803586. PMID 10772343. S2CID 8541209.
- Mori H, Okazawa H, Iwamoto K, Maeda E, Hashiramoto M, Kasuga M (Mar 2001). "A polymorphism in the 5' untranslated region and a Met229-->Leu variant in exon 5 of the human UCP1 gene are associated with susceptibility to type II diabetes mellitus". Diabetologia. 44 (3): 373–6. doi:10.1007/s001250051629. PMID 11317671.
- Nibbelink M, Moulin K, Arnaud E, Duval C, Pénicaud L, Casteilla L (Dec 2001). "Brown fat UCP1 is specifically expressed in uterine longitudinal smooth muscle cells". The Journal of Biological Chemistry. 276 (50): 47291–5. doi:10.1074/jbc.M105658200. PMID 11572862.
- Echtay KS, Roussel D, St-Pierre J, Jekabsons MB, Cadenas S, Stuart JA, Harper JA, Roebuck SJ, Morrison A, Pickering S, Clapham JC, Brand MD (Jan 2002). "Superoxide activates mitochondrial uncoupling proteins". Nature. 415 (6867): 96–9. Bibcode:2002Natur.415...96E. doi:10.1038/415096a. PMID 11780125. S2CID 4349744.
- Rousset S, del Mar Gonzalez-Barroso M, Gelly C, Pecqueur C, Bouillaud F, Ricquier D, Cassard-Doulcier AM (May 2002). "A new polymorphic site located in the human UCP1 gene controls the in vitro binding of CREB-like factor". International Journal of Obesity and Related Metabolic Disorders. 26 (5): 735–8. doi:10.1038/sj.ijo.0801973. PMID 12032762.
- Rim JS, Kozak LP (Sep 2002). "Regulatory motifs for CREB-binding protein and Nfe2l2 transcription factors in the upstream enhancer of the mitochondrial uncoupling protein 1 gene". The Journal of Biological Chemistry. 277 (37): 34589–600. doi:10.1074/jbc.M108866200. PMID 12084707.
- Kieć-Wilk B, Wybrańska I, Malczewska-Malec M, Leszczyńska-Gołabek L, Partyka L, Niedbał S, Jabrocka A, Dembińska-Kieć A (Sep 2002). "Correlation of the -3826A >G polymorphism in the promoter of the uncoupling protein 1 gene with obesity and metabolic disorders in obese families from southern Poland". Journal of Physiology and Pharmacology. 53 (3): 477–90. PMID 12375583.
External links
- Seaweed anti-obesity tablet hope (BBC - Thermogenin mentioned as part of process)
- thermogenin at the U.S. National Library of Medicine Medical Subject Headings (MeSH)