
Optogenetics is a biological technique to control the activity of neurons or other cell types with light. This is achieved by expression of light-sensitive ion channels, pumps or enzymes specifically in the target cells. On the level of individual cells, light-activated enzymes and transcription factors allow precise control of biochemical signaling pathways. In systems neuroscience, the ability to control the activity of a genetically defined set of neurons has been used to understand their contribution to decision making, learning, fear memory, mating, addiction, feeding, and locomotion. In a first medical application of optogenetic technology, vision was partially restored in a blind patient.
Optogenetic techniques have also been introduced to map the functional connectivity of the brain. By altering the activity of genetically labelled neurons with light and using imaging and electrophysiology techniques to record the activity of other cells, researchers can identify the statistical dependencies between cells and brain regions.
In a broader sense, optogenetics also includes methods to record cellular activity with genetically encoded indicators.
In 2010, optogenetics was chosen as the "Method of the Year" across all fields of science and engineering by the interdisciplinary research journal Nature Methods. At the same time, optogenetics was highlighted in the article on "Breakthroughs of the Decade" in the academic research journal Science.
History
In 1979, Francis Crick suggested that controlling all cells of one type in the brain, while leaving the others more or less unaltered, is a real challenge for neuroscience. Francis Crick speculated that a technology using light might be useful to control neuronal activity with temporal and spatial precision but at the time there was no technique to make neurons responsive to light.
By early 1990s LC Katz and E Callaway had shown that light could uncage glutamate. Heberle and Büldt in 1994 had already shown functional heterologous expression of a bacteriorhodopsin for light-activated ion flow in yeast. Later in 1995, Georg Nagel et al. and Ernst Bamberg tried the heterologous expression of microbial rhodopsins (also bacteriorhodopsin and also in a non-neural system, Xenopus oocytes) (Nagel et al., 1995, FEBS Lett.) and showed light-induced current.
The earliest genetically targeted method that used light to control rhodopsin-sensitized neurons was reported in January 2002, by Boris Zemelman and Gero Miesenböck, who employed Drosophila rhodopsin cultured mammalian neurons. In 2003, Zemelman and Miesenböck developed a second method for light-dependent activation of neurons in which single ionotropic channels TRPV1, TRPM8 and P2X2 were gated by photocaged ligands in response to light. Beginning in 2004, the Kramer and Isacoff groups developed organic photoswitches or "reversibly caged" compounds in collaboration with the Trauner group that could interact with genetically introduced ion channels. TRPV1 methodology, albeit without the illumination trigger, was subsequently used by several laboratories to alter feeding, locomotion and behavioral resilience in laboratory animals. However, light-based approaches for altering neuronal activity were not applied outside the original laboratories, likely because the easier to employ channelrhodopsin was cloned soon thereafter.
Peter Hegemann, studying the light response of green algae at the University of Regensburg, had discovered photocurrents that were too fast to be explained by the classic g-protein-coupled animal rhodopsins. Teaming up with the electrophysiologist Georg Nagel at the Max Planck Institute in Frankfurt, they could demonstrate that a single gene from the alga Chlamydomonas produced large photocurrents when expressed in the oocyte of a frog. To identify expressing cells, they replaced the cytoplasmic tail of the algal protein with a fluorescent protein YFP, generating the first generally applicable optogenetic tool. They stated in the 2003 paper that "expression of ChR2 in oocytes or mammalian cells may be used as a powerful tool to increase cytoplasmic Ca2+ concentration or to depolarize the cell membrane, simply by illumination".
Karl Deisseroth in the Bioengineering Department at Stanford published the notebook pages from early July 2004 of his initial experiment showing light activation of neurons expressing a channelrhodopsin. In August 2005, his laboratory staff, including graduate students Ed Boyden and Feng Zhang, in collaboration with Georg Nagel, published the first demonstration of a single-component optogenetic system, in neurons using the channelrhodopsin-2(H134R)-eYFP construct from Nagel and Hegemann.
Zhuo-Hua Pan of Wayne State University, researching on restore sight to blindness, tried channelrhodopsin out in ganglion cells—the neurons in our eyes that connect directly to the brain. Pan's first observation of optical activation of retinal neurons with channelrhodopsin was in August 2004 according to Pan, a month after Deisseroth's initial observation. Indeed, the transfected neurons became electrically active in response to light, and in 2005 Zhuo-Hua Pan reported successful in-vivo transfection of channelrhodopsin in retinal ganglion cells of mice, and electrical responses to photostimulation in retinal slice culture. This approach was eventually realized in a human patient by Botond Roska and coworkers in 2021.
In April 2005, Susana Lima and Miesenböck reported the first use of genetically-targeted P2X2 photostimulation to control the behaviour of an animal. They showed that photostimulation of genetically circumscribed groups of neurons, such as those of the dopaminergic system, elicited characteristic behavioural changes in fruit flies.
In October 2005, Lynn Landmesser and Stefan Herlitze also published the use of channelrohodpsin-2 to control neuronal activity in cultured hippocampal neurons and chicken spinal cord circuits in intact developing embryos. In addition, they introduced for the first time vertebrate rhodopsin, a light-activated G protein coupled receptor, as a tool to inhibit neuronal activity via the recruitment of intracellular signaling pathways also in hippocampal neurons and the intact developing chicken embryo.
The groups of Alexander Gottschalk and Georg Nagel made the first ChR2 mutant (H134R) and were first to use channelrhodopsin-2 for controlling neuronal activity in an intact animal, showing that motor patterns in the roundworm C. elegans could be evoked by light stimulation of genetically selected neural circuits (published in December 2005). In mice, controlled expression of optogenetic tools is often achieved with cell-type-specific Cre/loxP methods developed for neuroscience by Joe Z. Tsien back in the 1990s to activate or inhibit specific brain regions and cell-types in vivo.
In 2007, the labs of Boyden and Deisseroth (together with the groups of Gottschalk and Nagel) simultaneously reported successful optogenetic inhibition of activity in neurons.
In 2007, Nagel and Hegemann's groups started the optogenetic manipulation of cAMP. In 2014, Avelar et al. reported the first rhodopsin-guanylyl cyclase gene from fungus. In 2015, Scheib et al. and Gao et al. characterized the activity of the rhodopsin-guanylyl cyclase gene. And Shiqiang Gao et al. and Georg Nagel, Alexander Gottschalk identified it as the first 8 TM enzyme rhodopsin.
Awards
The powerful impact of optogenetic technology on brain research has been recognized by numerous awards to key players in the field.
In 2010, Georg Nagel, Peter Hegemann and Ernst Bamberg were awarded the Wiley Prize in Biomedical Sciences and they were also among those awarded the Karl Heinz Beckurts Prize in 2010. In the same year, Karl Deisseroth was awarded the inaugural HFSP Nakasone Award for "his pioneering work on the development of optogenetic methods for studying the function of neuronal networks underlying behavior".
In 2012, Bamberg, Deisseroth, Hegemann and Nagel were awarded the Zülch Prize by the Max Planck Society, and Miesenböck was awarded the Baillet Latour Health Prize for "having pioneered optogenetic approaches to manipulate neuronal activity and to control animal behaviour."
In 2013, Nagel and Hegemann were among those awarded the Louis-Jeantet Prize for Medicine. Also that year, year Bamberg, Boyden, Deisseroth, Hegemann, Miesenböck and Nagel were jointly awarded The Brain Prize for "their invention and refinement of optogenetics."
In 2017, Deisseroth was awarded the Else Kröner Fresenius Research Prize for "his discoveries in optogenetics and hydrogel-tissue chemistry, as well as his research into the neural circuit basis of depression."
In 2018, the Inamori Foundation presented Deisseroth with the Kyoto Prize for "spearheading optogenetics” and "revolutionizing systems neuroscience research."
In 2019, Bamberg, Boyden, Deisseroth, Hegemann, Miesenböck and Nagel were awarded the Rumford Prize by the American Academy of Arts and Sciences in recognition of "their extraordinary contributions related to the invention and refinement of optogenetics."
In 2020, Deisseroth was awarded the Heineken Prize for Medicine from the Royal Netherlands Academy of Arts and Sciences, for developing optogenetics and hydrogel-tissue chemistry. On the same year, Miesenböck, Hegemann and Nagel jointly received the Shaw Prize in Life Science and Medicine.
In 2021, Hegemann, Deisseroth and Dieter Oesterhelt received the Albert Lasker Award for Basic Medical Research.
Description
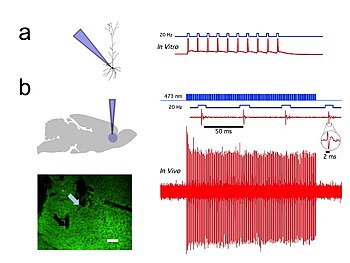
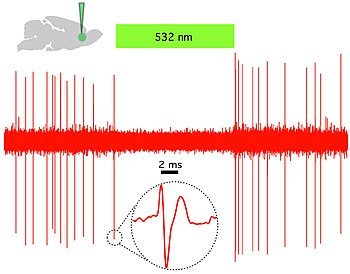
Optogenetics provides millisecond-scale temporal precision which allows the experimenter to keep pace with fast biological information processing (for example, in probing the causal role of specific action potential patterns in defined neurons). Indeed, to probe the neural code, optogenetics by definition must operate on the millisecond timescale to allow addition or deletion of precise activity patterns within specific cells in the brains of intact animals, including mammals (see Figure 1). By comparison, the temporal precision of traditional genetic manipulations (employed to probe the causal role of specific genes within cells, via "loss-of-function" or "gain of function" changes in these genes) is rather slow, from hours or days to months. It is important to also have fast readouts in optogenetics that can keep pace with the optical control. This can be done with electrical recordings ("optrodes") or with reporter proteins that are biosensors, where scientists have fused fluorescent proteins to detector proteins. Additionally, beyond its scientific impact optogenetics represents an important case study in the value of both ecological conservation (as many of the key tools of optogenetics arise from microbial organisms occupying specialized environmental niches), and in the importance of pure basic science as these opsins were studied over decades for their own sake by biophysicists and microbiologists, without involving consideration of their potential value in delivering insights into neuroscience and neuropsychiatric disease.
Light-activated proteins: channels, pumps and enzymes
The hallmark of optogenetics therefore is introduction of fast light-activated channels, pumps, and enzymes that allow temporally precise manipulation of electrical and biochemical events while maintaining cell-type resolution through the use of specific targeting mechanisms. Among the microbial opsins which can be used to investigate the function of neural systems are the channelrhodopsins (ChR2, ChR1, VChR1, and SFOs) to excite neurons and anion-conducting channelrhodopsins for light-induced inhibition. Indirectly light-controlled potassium channels have recently been engineered to prevent action potential generation in neurons during blue light illumination. Light-driven ion pumps are also used to inhibit neuronal activity, e.g. halorhodopsin (NpHR), enhanced halorhodopsins (eNpHR2.0 and eNpHR3.0, see Figure 2),archaerhodopsin (Arch), fungal opsins (Mac) and enhanced bacteriorhodopsin (eBR).
Optogenetic control of well-defined biochemical events within behaving mammals is now also possible. Building on prior work fusing vertebrate opsins to specific G-protein coupled receptors a family of chimeric single-component optogenetic tools was created that allowed researchers to manipulate within behaving mammals the concentration of defined intracellular messengers such as cAMP and IP3 in targeted cells. Other biochemical approaches to optogenetics (crucially, with tools that displayed low activity in the dark) followed soon thereafter, when optical control over small GTPases and adenylyl cyclase was achieved in cultured cells using novel strategies from several different laboratories.Photoactivated adenylyl cyclases have been discovered in fungi and successfully used to control cAMP levels in mammalian neurons. This emerging repertoire of optogenetic actuators now allows cell-type-specific and temporally precise control of multiple axes of cellular function within intact animals.
Hardware for light application
Another necessary factor is hardware (e.g. integrated fiberoptic and solid-state light sources) to allow specific cell types, even deep within the brain, to be controlled in freely behaving animals. Most commonly, the latter is now achieved using the fiberoptic-coupled diode technology introduced in 2007, though to avoid use of implanted electrodes, researchers have engineered ways to inscribe a "window" made of zirconia that has been modified to be transparent and implanted in mice skulls, to allow optical waves to penetrate more deeply to stimulate or inhibit individual neurons. To stimulate superficial brain areas such as the cerebral cortex, optical fibers or LEDs can be directly mounted to the skull of the animal. More deeply implanted optical fibers have been used to deliver light to deeper brain areas. Complementary to fiber-tethered approaches, completely wireless techniques have been developed utilizing wirelessly delivered power to headborne LEDs for unhindered study of complex behaviors in freely behaving organisms.
Expression of optogenetic actuators
Optogenetics also necessarily includes the development of genetic targeting strategies such as cell-specific promoters or other customized conditionally-active viruses, to deliver the light-sensitive probes to specific populations of neurons in the brain of living animals (e.g. worms, fruit flies, mice, rats, and monkeys). In invertebrates such as worms and fruit flies some amount of all-trans-retinal (ATR) is supplemented with food. A key advantage of microbial opsins as noted above is that they are fully functional without the addition of exogenous co-factors in vertebrates.
Technique
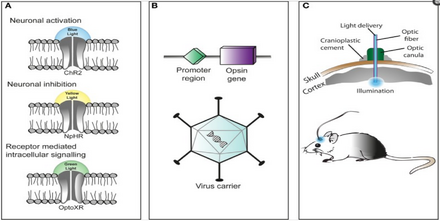
The technique of using optogenetics is flexible and adaptable to the experimenter's needs. Cation-selective channelrhodopsins (e.g. ChR2) are used to excite neurons, anion-conducting channelrhodopsins (e.g. GtACR2) inhibit neuronal activity. Combining these tools into a single construct (e.g. BiPOLES) allows for both inhibition and excitation, depending on the wavelength of illumination.
Introducing the microbial opsin into a specific subset of cells is challenging. A popular approach is to introduce an engineered viral vector that contains the optogenetic actuator gene attached to a specific promoter such as CAMKIIα, which is active in excitatory neurons. This allows for some level of specificity, preventing e.g. expression in glia cells.
A more specific approach is based on transgenic "driver" mice which express Cre recombinase, an enzyme that catalyzes recombination between two lox-P sites, in a specific subset of cells, e.g. parvalbumin-expressing interneurons. By introducing an engineered viral vector containing the optogenetic actuator gene in between two lox-P sites, only the cells producing Cre recombinase will express the microbial opsin. This technique has allowed for multiple modified optogenetic actuators to be used without the need to create a whole line of transgenic animals every time a new microbial opsin is needed.
After the introduction and expression of the microbial opsin, a computer-controlled light source has to be optically coupled to the brain region in question. Light-emitting diodes (LEDs) or fiber-coupled diode-pumped solid-state lasers (DPSS) are frequently used. Recent advances include the advent of wireless head-mounted devices that apply LEDs to the targeted areas and as a result, give the animals more freedom to move.
Fiber-based approaches can also be used to combine optical stimulation and calcium imaging. This enables researchers to visualize and manipulate the activity of single neurons in awake behaving animals. It is also possible to record from multiple deep brain regions at the same using GRIN lenses connected via optical fiber to an externally positioned photodetector and photostimulator.
Technical challenges
Selective expression
One of the main problems of optogenetics is that not all the cells in question may express the microbial opsin gene at the same level. Thus, even illumination with a defined light intensity will have variable effects on individual cells. Optogenetic stimulation of neurons in the brain is even less controlled as the light intensity drops exponentially from the light source (e.g. implanted optical fiber).
It remains difficult to target opsin to defined subcellular compartments, e.g. the plasma membrane, synaptic vesicles, or mitochondria. Restricting the opsin to specific regions of the plasma membrane such as dendrites, somata or axon terminals provides a more robust understanding of neuronal circuitry.
Mathematical modelling shows that selective expression of opsin in specific cell types can dramatically alter the dynamical behavior of the neural circuitry. In particular, optogenetic stimulation that preferentially targets inhibitory cells can transform the excitability of the neural tissue, affecting non-transfected neurons as well.
Kinetics and synchronization
The original channelrhodopsin-2 was slower closing than typical cation channels of cortical neurons, leading to prolonged depolarization and calcium influx. Many channelrhodopsin variants with more favorable kinetics have since been engineered.[55][56]
A difference between natural spike patterns and optogenetic activation is that pulsed light stimulation produces synchronous activation of expressing neurons, which removes the possibility of sequential activity in the stimulated population. Therefore, it is difficult to understand how the cells in the population affected communicate with one another or how their phasic properties of activation relate to circuit function.
Optogenetic activation has been combined with functional magnetic resonance imaging (ofMRI) to elucidate the connectome, a thorough map of the brain's neural connections. Precisely timed optogenetic activation is used to calibrate the delayed hemodynamic signal (BOLD) fMRI is based on.
Light absorption spectrum
The opsin proteins currently in use have absorption peaks across the visual spectrum, but remain considerably sensitive to blue light. This spectral overlap makes it very difficult to combine opsin activation with genetically encoded indicators (GEVIs, GECIs, GluSnFR, synapto-pHluorin), most of which need blue light excitation. Opsins with infrared activation would, at a standard irradiance value, increase light penetration and augment resolution through reduction of light scattering.
Spatial response
Due to scattering, a narrow light beam to stimulate neurons in a patch of neural tissue can evoke a response profile that is much broader than the stimulation beam. In this case, neurons may be activated (or inhibited) unintentionally. Computational simulation tools are used to estimate the volume of stimulated tissue for different wavelengths of light.
Applications
The field of optogenetics has furthered the fundamental scientific understanding of how specific cell types contribute to the function of biological tissues such as neural circuits in vivo. On the clinical side, optogenetics-driven research has led to insights into Parkinson's disease and other neurological and psychiatric disorders such as autism, Schizophrenia, drug abuse, anxiety, and depression. An experimental treatment for blindness involves a channel rhodopsin expressed in ganglion cells, stimulated with light patterns from engineered goggles.
Identification of particular neurons and networks
Amygdala
Optogenetic approaches have been used to map neural circuits in the amygdala that contribute to fear conditioning. One such example of a neural circuit is the connection made from the basolateral amygdala to the dorsal-medial prefrontal cortex where neuronal oscillations of 4 Hz have been observed in correlation to fear induced freezing behaviors in mice. Transgenic mice were introduced with channelrhodoposin-2 attached with a parvalbumin-Cre promoter that selectively infected interneurons located both in the basolateral amygdala and the dorsal-medial prefrontal cortex responsible for the 4 Hz oscillations. The interneurons were optically stimulated generating a freezing behavior and as a result provided evidence that these 4 Hz oscillations may be responsible for the basic fear response produced by the neuronal populations along the dorsal-medial prefrontal cortex and basolateral amygdala.
Olfactory bulb
Optogenetic activation of olfactory sensory neurons was critical for demonstrating timing in odor processing and for mechanism of neuromodulatory mediated olfactory guided behaviors (e.g. aggression, mating) In addition, with the aid of optogenetics, evidence has been reproduced to show that the "afterimage" of odors is concentrated more centrally around the olfactory bulb rather than on the periphery where the olfactory receptor neurons would be located. Transgenic mice infected with channel-rhodopsin Thy1-ChR2, were stimulated with a 473 nm laser transcranially positioned over the dorsal section of the olfactory bulb. Longer photostimulation of mitral cells in the olfactory bulb led to observations of longer lasting neuronal activity in the region after the photostimulation had ceased, meaning the olfactory sensory system is able to undergo long term changes and recognize differences between old and new odors.
Nucleus accumbens
Optogenetics, freely moving mammalian behavior, in vivo electrophysiology, and slice physiology have been integrated to probe the cholinergic interneurons of the nucleus accumbens by direct excitation or inhibition. Despite representing less than 1% of the total population of accumbal neurons, these cholinergic cells are able to control the activity of the dopaminergic terminals that innervate medium spiny neurons (MSNs) in the nucleus accumbens. These accumbal MSNs are known to be involved in the neural pathway through which cocaine exerts its effects, because decreasing cocaine-induced changes in the activity of these neurons has been shown to inhibit cocaine conditioning. The few cholinergic neurons present in the nucleus accumbens may prove viable targets for pharmacotherapy in the treatment of cocaine dependence
Prefrontal cortex

In vivo and in vitro recordings from the University of Colorado, Boulder Optophysiology Laboratory of Donald C. Cooper Ph.D. showing individual CAMKII AAV-ChR2 expressing pyramidal neurons within the prefrontal cortex that demonstrated high fidelity action potential output with short pulses of blue light at 20 Hz (Figure 1).
Motor cortex
In vivo repeated optogenetic stimulation in healthy animals was able to eventually induce seizures. This model has been termed optokindling.
Piriform cortex
In vivo repeated optogenetic stimulation of pyramidal cells of the piriform cortex in healthy animals was able to eventually induce seizures.In vitro studies have revealed a loss of feedback inhibition in the piriform circuit due to impaired GABA synthesis.
Heart
Optogenetics was applied on atrial cardiomyocytes to end spiral wave arrhythmias, found to occur in atrial fibrillation, with light. This method is still in the development stage. A recent study explored the possibilities of optogenetics as a method to correct for arrythmias and resynchronize cardiac pacing. The study introduced channelrhodopsin-2 into cardiomyocytes in ventricular areas of hearts of transgenic mice and performed in vitro studies of photostimulation on both open-cavity and closed-cavity mice. Photostimulation led to increased activation of cells and thus increased ventricular contractions resulting in increasing heart rates. In addition, this approach has been applied in cardiac resynchronization therapy (CRT) as a new biological pacemaker as a substitute for electrode based-CRT. Lately, optogenetics has been used in the heart to defibrillate ventricular arrhythmias with local epicardial illumination, a generalized whole heart illumination or with customized stimulation patterns based on arrhythmogenic mechanisms in order to lower defibrillation energy.
Spiral ganglion
Optogenetic stimulation of the spiral ganglion in deaf mice restored auditory activity. Optogenetic application onto the cochlear region allows for the stimulation or inhibition of the spiral ganglion cells (SGN). In addition, due to the characteristics of the resting potentials of SGN's, different variants of the protein channelrhodopsin-2 have been employed such as Chronos, CatCh and f-Chrimson. Chronos and CatCh variants are particularly useful in that they have less time spent in their deactivated states, which allow for more activity with less bursts of blue light emitted. Additionally, using engineered red-shifted channels as f-Chrimson allow for stimulation using longer wavelengths, which decreases the potential risks of phototoxicity in the long term without compromising gating speed. The result being that the LED producing the light would require less energy and the idea of cochlear prosthetics in association with photo-stimulation, would be more feasible.
Brainstem
Optogenetic stimulation of a modified red-light excitable channelrhodopsin (ReaChR) expressed in the facial motor nucleus enabled minimally invasive activation of motoneurons effective in driving whisker movements in mice. One novel study employed optogenetics on the Dorsal Raphe Nucleus to both activate and inhibit dopaminergic release onto the ventral tegmental area. To produce activation transgenic mice were infected with channelrhodopsin-2 with a TH-Cre promoter and to produce inhibition the hyperpolarizing opsin NpHR was added onto the TH-Cre promoter. Results showed that optically activating dopaminergic neurons led to an increase in social interactions, and their inhibition decreased the need to socialize only after a period of isolation.
Visual system
Studying the visual system using optogenetics can be challenging. Indeed, the light used for optogenetic control may lead to the activation of photoreceptors, as a result of the proximity between primary visual circuits and these photoreceptors. In this case, spatial selectivity is difficult to achieve (particularly in the case of the fly optic lobe). Thus, the study of the visual system requires spectral separation, using channels that are activated by different wavelengths of light than rhodopsins within the photoreceptors (peak activation at 480 nm for Rhodopsin 1 in Drosophila). Red-shifted CsChrimson or bistable Channelrhodopsin are used for optogenetic activation of neurons (i.e. depolarization), as both allow spectral separation. In order to achieve neuronal silencing (i.e. hyperpolarization), an anion channelrhodopsin discovered in the cryptophyte algae species Guillardia theta (named GtACR1). can be used. GtACR1 is more light sensitive than other inhibitory channels such as the Halorhodopsin class of chlorid pumps and imparts a strong conductance. As its activation peak (515 nm) is close to that of Rhodopsin 1, it is necessary to carefully calibrate the optogenetic illumination as well as the visual stimulus. The factors to take into account are the wavelength of the optogenetic illumination (possibly higher than the activation peak of GtACR1), the size of the stimulus (in order to avoid the activation of the channels by the stimulus light) and the intensity of the optogenetic illumination. It has been shown that GtACR1 can be a useful inhibitory tool in optogenetic study of Drosophila's visual system by silencing T4/T5 neurons expression. These studies can also be led on intact behaving animals, for instance to probe optomotor response.
Sensorimotor system
Optogenetically inhibiting or activating neurons tests their necessity and sufficiency, respectively, in generating a behavior. Using this approach, researchers can dissect the neural circuitry controlling motor output. By perturbing neurons at various places in the sensorimotor system, researchers have learned about the role of descending neurons in eliciting stereotyped behaviors, how localized tactile sensory input and activity of interneurons alters locomotion, and the role of Purkinje cells in generating and modulating movement. This is a powerful technique for understanding the neural underpinnings of animal locomotion and movement more broadly.
Precise temporal control of interventions
The currently available optogenetic actuators allow for the accurate temporal control of the required intervention (i.e. inhibition or excitation of the target neurons) with precision routinely going down to the millisecond level. The temporal precision varies, however, across optogenetic actuators, and depends on the frequency and intensity of the stimulation.
Experiments can now be devised where the light used for the intervention is triggered by a particular element of behavior (to inhibit the behavior), a particular unconditioned stimulus (to associate something to that stimulus) or a particular oscillatory event in the brain (to inhibit the event). This kind of approach has already been used in several brain regions:
Hippocampus
Sharp waves and ripple complexes (SWRs) are distinct high frequency oscillatory events in the hippocampus thought to play a role in memory formation and consolidation. These events can be readily detected by following the oscillatory cycles of the on-line recorded local field potential. In this way the onset of the event can be used as a trigger signal for a light flash that is guided back into the hippocampus to inhibit neurons specifically during the SWRs and also to optogenetically inhibit the oscillation itself. These kinds of "closed-loop" experiments are useful to study SWR complexes and their role in memory.
Cellular biology/cell signaling pathways
Analogously to how natural light-gated ion channels such as channelrhodopsin-2 allows optical control of ion flux, which is especially useful in neuroscience, natural light-controlled signal transduction proteins also allow optical control of biochemical pathways, including both second-messenger generation and protein-protein interactions, which is especially useful in studying cell and developmental biology. In 2002, the first example of using photoproteins from another organism for controlling a biochemical pathway was demonstrated using the light-induced interaction between plant phytochrome and phytochrome-interacting factor (PIF) to control gene transcription in yeast. By fusing phytochrome to a DNA-binding domain and PIF to a transcriptional activation domain, transcriptional activation of genes recognized by the DNA-binding domain could be induced by light. This study anticipated aspects of the later development of optogenetics in the brain, for example, by suggesting that "Directed light delivery by fiber optics has the potential to target selected cells or tissues, even within larger, more-opaque organisms." The literature has been inconsistent as to whether control of cellular biochemistry with photoproteins should be subsumed within the definition of optogenetics, as optogenetics in common usage refers specifically to the control of neuronal firing with opsins, and as control of neuronal firing with opsins postdates and utilizes distinct mechanisms from control of cellular biochemistry with photoproteins.
Photosensitive proteins utilized in various cell signaling pathways
In addition to phytochromes, which are found in plants and cyanobacteria, LOV domains(Light-oxygen-voltage-sensing domain) from plants and yeast and cryptochrome domains from plants are other natural photosensory domains that have been used for optical control of biochemical pathways in cells. In addition, a synthetic photosensory domain has been engineered from the fluorescent protein Dronpa for optical control of biochemical pathways. In photosensory domains, light absorption is either coupled to a change in protein-protein interactions (in the case of phytochromes, some LOV domains, cryptochromes, and Dronpa mutants) or a conformational change that exposes a linked protein segment or alters the activity of a linked protein domain (in the case of phytochromes and some LOV domains). Light-regulated protein-protein interactions can then be used to recruit proteins to DNA, for example to induce gene transcription or DNA modifications, or to the plasma membrane, for example to activate resident signaling proteins. CRY2 also clusters when active, so has been fused with signaling domains and subsequently photoactivated to allow for clustering-based activation. The LOV2 domain of Avena sativa(common oat) has been used to expose short peptides or an active protein domain in a light-dependent manner. Introduction of this LOV domain into another protein can regulate function through light induced peptide disorder. The asLOV2 protein, which optogenetically exposes a peptide, has also been used as a scaffold for several synthetic light induced dimerization and light induced dissociation systems (iLID and LOVTRAP, respectively). The systems can be used to control proteins through a protein splitting strategy. Photodissociable Dronpa domains have also been used to cage a protein active site in the dark, uncage it after cyan light illumination, and recage it after violet light illumination.
Temporal control of signal transduction with light
The ability to optically control signals for various time durations is being explored to elucidate how cell signaling pathways convert signal duration and response to different outputs. Natural signaling cascades are capable of responding with different outputs to differences in stimulus timing duration and dynamics. For example, treating PC12 cells with epidermal growth factor (EGF, inducing a transient profile of ERK activity) leads to cellular proliferation whereas introduction of nerve growth factor (NGF, inducing a sustained profile of ERK activity) leads to differentiation into neuron-like cells. This behavior was initially characterized using EGF and NGF application, but the finding has been partially replicated with optical inputs. In addition, a rapid negative feedback loop in the RAF-MEK-ERK pathway was discovered using pulsatile activation of a photoswitchable RAF engineered with photodissociable Dronpa domains.
Optogenetic noise-photostimulation
Professor Elias Manjarrez´s research group introduced the Optogenetic noise-photostimulation. This is a technique that uses random noisy light to activate neurons expressing ChR2. An optimal level of optogenetic-noise photostimulation on the brain can increase the somatosensory evoked field potentials, the firing frequency response of pyramidal neurons to somatosensory stimulation, and the sodium current amplitude.
Further reading
- Appasani K (2017). Optogenetics: from neuronal function to mapping and disease biology. Cambridge, UK: Cambridge University Press. ISBN 978-1-107-05301-4.
- Banerjee S, Mitra D (January 2020). "Structural Basis of Design and Engineering for Advanced Plant Optogenetics". Trends in Plant Science. 25 (1): 35–65. doi:10.1016/j.tplants.2019.10.002. PMID 31699521. S2CID 207942668.
- Hu W, Li Q, Li B, Ma K, Zhang C, Fu X (January 2020). "Optogenetics sheds new light on tissue engineering and regenerative medicine". Biomaterials. 227: 119546. doi:10.1016/j.biomaterials.2019.119546. PMID 31655444. S2CID 204918731.
- Jarrin S, Finn DP (October 2019). "Optogenetics and its application in pain and anxiety research". Neuroscience and Biobehavioral Reviews. 105: 200–211. doi:10.1016/j.neubiorev.2019.08.007. PMID 31421140. S2CID 199577276.
- Johnson HE, Toettcher JE (August 2018). "Illuminating developmental biology with cellular optogenetics". Current Opinion in Biotechnology. 52: 42–48. doi:10.1016/j.copbio.2018.02.003. PMC 6082700. PMID 29505976.
- Krueger D, Izquierdo E, Viswanathan R, Hartmann J, Pallares Cartes C, De Renzis S (October 2019). "Principles and applications of optogenetics in developmental biology". Development. 146 (20): dev175067. doi:10.1242/dev.175067. PMC 6914371. PMID 31641044.
- Losi A, Gardner KH, Möglich A (November 2018). "Blue-Light Receptors for Optogenetics". Chemical Reviews. 118 (21): 10659–10709. doi:10.1021/acs.chemrev.8b00163. PMC 6500593. PMID 29984995.
- Vriz S, Ozawa T (September 2018). Optogenetics: light-driven actuators and light-emitting sensors in cell biology. Comprehensive Series in Photochemistry and Photobiology. Vol. 18. London: Royal Society of Chemistry. ISBN 978-1-78801-237-9.
- Wittmann T, Dema A, van Haren J (October 2020). "Lights, cytoskeleton, action: Optogenetic control of cell dynamics". Current Opinion in Cell Biology. Elsevier Ltd. 66: 1–10. doi:10.1016/j.ceb.2020.03.003. PMC 7577957. PMID 32371345.
External links
- "Optogenetics: shedding light on the brain's secrets". Scientifica.
- "Optogenetics: Integrated Calcium Imaging and Optogenetics". Inscopix.
| Optogenetic actuators | |
|---|---|
| Optogenetic sensors | |
| Related techniques | |
| Technologies |  |
|
|---|---|---|
| Scientific phenomena | ||
| Disciplines | ||
| Speculative | ||
| People | ||
| Other | ||