
Eukaryogenesis, the process which created the eukaryotic cell and lineage, is a milestone in the evolution of life, since eukaryotes include all complex cells and almost all multicellular organisms. The process is widely agreed to have involved symbiogenesis, in which archaea and bacteria came together to create the first eukaryotic common ancestor (FECA). This cell had a new level of complexity and capability, with a nucleus, at least one centriole and cilium, facultatively aerobic mitochondria, sex (meiosis and syngamy), a dormant cyst with a cell wall of chitin and/or cellulose and peroxisomes. It evolved into a population of single-celled organisms that included the last eukaryotic common ancestor (LECA), gaining capabilities along the way, though the sequence of the steps involved has been disputed, and may not have started with symbiogenesis. In turn, the LECA gave rise to the eukaryotes' crown group, containing the ancestors of animals, fungi, plants, and a diverse range of single-celled organisms.
Context
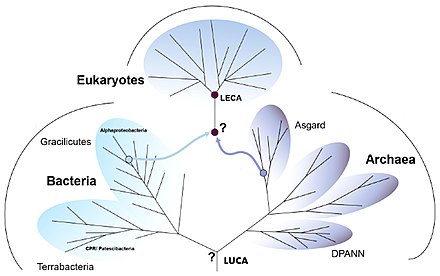
Life arose on Earth once it had cooled enough for oceans to form. The last universal common ancestor (LUCA) was an organism which had ribosomes and the genetic code; it lived some 4 billion years ago. It gave rise to two main branches of prokaryotic life, the bacteria and the archaea. From among these small-celled, rapidly-dividing ancestors arose the Eukaryotes, with much larger cells, nuclei, and distinctive biochemistry. The eukaryotes form a domain that contains all complex cells and most types of multicellular organism, including the animals, plants, and fungi.
Symbiogenesis

According to the theory of symbiogenesis (also known as the endosymbiotic theory) championed by Lynn Margulis, a member of the archaea gained a bacterial cell as a component. The archaeal cell was a member of the Asgard group. The bacterium was one of the Alphaproteobacteria, which had the ability to use oxygen in its respiration. This enabled it – and the archaeal cells that included it – to survive in the presence of oxygen, which was poisonous to other organisms adapted to reducing conditions. The endosymbiotic bacteria became the eukaryotic cell's mitochondria, providing most of the energy of the cell.Lynn Margulis and colleagues have suggested that the cell also acquired a Spirochaete bacterium as a symbiont, providing the cell skeleton of microtubules and the ability to move, including the ability to pull chromosomes into two sets during mitosis, cell division. More recently, the Asgard archaean has been identified as belonging to the Heimdallarchaeota.
Last eukaryotic common ancestor (LECA)
The last eukaryotic common ancestor (LECA) is the hypothetical last common ancestor of all living eukaryotes, around 2 gya and was most likely a biological population. It is believed to have been a protist with a nucleus, at least one centriole and cilium, facultatively aerobic mitochondria, sex (meiosis and syngamy), a dormant cyst with a cell wall of chitin and/or cellulose, and peroxisomes.
It had been proposed that the LECA fed by phagocytosis, engulfing other organisms. However, in 2022, Nico Bremer and colleagues confirmed that the LECA had mitochondria, and stated that it had multiple nuclei, but disputed that it was phagotrophic, meaning that the ability found in many eukaryotes to engulf materials developed later, rather than being acquired first and then being used to engulf the alphaproteobacteria that became mitochondria.
The LECA has been described as having "spectacular cellular complexity". Its cell was divided into compartments. It appears to have inherited a set of ensosomal sorting complex (ESCRT) proteins that enable membranes to be remodelled, including pinching off vesicles to form endosomes. Its apparatus for transcribing DNA into RNA, and then translating the RNA into proteins, were separated, allowing the expression of genes to become more complex. It had mechanisms for reshuffling its genetic material and possibly manipulating its own evolvability. All of these gave the LECA "a compelling cohort of selective advantages".
Scenarios
Biologists have proposed multiple scenarios for the creation of the eukaryotes. While there is broad agreement that the LECA must have had a nucleus, mitochondria, and internal membranes, the order in which these were acquired has been disputed. In the syntrophic model, the first eukaryotic common ancestor (FECA, around 2.2 gya) gained mitochondria, then membranes, then a nucleus. In the phagotrophic model, it gained a nucleus, then membranes, then mitochondria. In a more complex process, it gained all three in short order, then other capabilities. Other models have been proposed. Whatever happened, many lineages must have been created, but the LECA either out-competed or came together with the other lineages to form a single point of origin for the eukaryotes.Nick Lane and William Martin have argued that mitochondria came first, on the grounds that energy had been the limiting factor on the size of the prokaryotic cell. The phagotrophic model presupposes the ability to engulf food, enabling the cell to engulf the aerobic bacterium that became the mitochondrion.
Eugene Koonin and others, noting that the archaea share many features with eukaryotes, argue that rudimentary eukaryotic traits such as membrane-lined compartments were acquired before endosymbiosis added mitochondria to the early eukaryotic cell, while the cell wall was lost. In the same way, mitochondrial acquisition must not be regarded as the end of the process, for still new complex families of genes had to be developed after or during the endosymbiotic exchange. In this way, from FECA to LECA, we can think of organisms that can be considered as protoeukaryotes. At the end of the process, LECA was already a complex organism with the presence of protein families involved in cellular compartmentalization.
Diversification: crown eukaryotes
In turn, the LECA gave rise to the eukaryotes' crown group, containing the ancestors of animals, fungi, plants, and a diverse range of single-celled organisms with the new capabilities and complexity of the eukaryotic cell. Single cells without cell walls are fragile and have a low probability of being fossilised. If fossilised, they have few features to distinguish them clearly from prokaryotes: size, morphological complexity, and (eventually) multicellularity. Early eukaryote fossils, from the late Paleoproterozoic, include acritarch microfossils with relatively robust ornate carbonaceous vesicles of Tappania from 1.63 gya and Shuiyousphaeridium from 1.8 gya.