
| Protist Temporal range:
| |
|---|---|

| |
|
Scientific classification | |
| Domain: | Eukaryota |
| Supergroups | |
|
Amoebozoa | |
| Cladistically included but traditionally excluded taxa | |
|
Animalia | |
A protist (/ˈproʊtɪst/ PROH-tist) is any eukaryotic organism (that is, an organism whose cells contain a cell nucleus) that is not an animal, plant, or fungus. Protists, along with other eukaryotes, all descend from the last eukaryotic common ancestor. Protists do not form a natural group, or clade; any unicellular eukaryote may be described as a protist, in addition to some cases of multicellular protists such as slime molds, brown algae and xenophyophorean forams. The study of protists is termed protistology.
Overview
Besides their relatively simple levels of organization, protists do not necessarily have much in common. When used, the term "protists" is now considered to mean a paraphyletic assemblage of similar-appearing but diverse taxa (biological groups); these taxa do not have an exclusive common ancestor beyond being composed of eukaryotes, and have different life cycles, trophic levels, modes of locomotion, and cellular structures.
Examples of protists include:
- Amoebas (including nucleariids and Foraminifera);
- Choanoflagellates; ciliates;
- Diatoms;
- Dinoflagellates;
- Giardia;
- Oomycetes (including Phytophthora, the cause of the Great Famine of Ireland); and
- Plasmodium (which causes malaria);
- slime molds.
These examples are unicellular, although oomycetes can join to form filaments, and slime molds can aggregate into a tissue-like mass.
In cladistic systems (classifications based on common ancestry), there are no equivalents to the taxa Protista or Protoctista, as both terms refer to a paraphyletic group that spans the entire eukaryotic branch of the tree of life. In cladistic classification, the contents of Protista are mostly distributed among various supergroups: examples include the
- Archaeplastida (or Plantae sensu lato);
- Excavata (which is mostly unicellular flagellates); and
- Opisthokonta (which commonly includes unicellular flagellates, but also animals and fungi).
- SAR supergroup (of stramenopiles or heterokonts, alveolates, and Rhizaria);
"Protista", "Protoctista", and "Protozoa" are therefore considered obsolete. However, the term "protist" continues to be used informally as a catch-all term for eukaryotic organisms that are not within other traditional kingdoms. For example, the word "protist pathogen" may be used to denote any disease-causing organism that is not plant, animal, fungal, prokaryotic, viral, or subviral.
Subdivisions
The term Protista was first used by Ernst Haeckel in 1866. Protists were traditionally subdivided into several groups based on similarities to the "higher" kingdoms such as:
- Protozoa
- Protozoans are unicellular "animal-like" (heterotrophic, and sometimes parasitic) organisms that are further sub-divided based on characteristics such as motility, such as the (flagellated) Flagellata, the (ciliated) Ciliophora, the (phagocytic) amoeba, and the (spore-forming) Sporozoa.
- Protophyta
- Protophyta are "plant-like" (autotrophic) organisms that are composed mostly of unicellular algae. The dinoflagellates, diatoms and Euglena-like flagellates are photosynthetic protists.
- Mold
- Molds generally refer to fungi; but slime molds and water molds are "fungus-like" (saprophytic) protists, although some are pathogens. Two separate types of slime molds exist, the cellular and acellular forms.
Some protists, sometimes called ambiregnal protists, have been considered to be both protozoa and algae or fungi (e.g., slime molds and flagellated algae), and names for these have been published under either or both of the ICN and the ICZN. Conflicts, such as these – for example the dual-classification of Euglenids and Dinobryons, which are mixotrophic – is an example of why the kingdom Protista was adopted.
These traditional subdivisions, largely based on superficial commonalities, have been replaced by classifications based on phylogenetics (evolutionary relatedness among organisms). Molecular analyses in modern taxonomy have been used to redistribute former members of this group into diverse and sometimes distantly related phyla. For instance, the water molds are now considered to be closely related to photosynthetic organisms such as Brown algae and Diatoms, the slime molds are grouped mainly under Amoebozoa, and the Amoebozoa itself includes only a subset of the "Amoeba" group, and significant number of erstwhile "Amoeboid" genera are distributed among Rhizarians and other Phyla.
However, the older terms are still used as informal names to describe the morphology and ecology of various protists. For example, the term protozoa is used to refer to heterotrophic species of protists that do not form filaments.
History of classification
Early concepts of protists

The father of protistology, Anton van Leeuwenhoek, is thought to be the first person to observe a variety of free-living protists, which he referred to as “very little animalcules” in 1674.
From the start of the 18th century, the popular term “infusion animals” was introduced by Ledermuller in 1763 to refer to these small organisms, and was later formalized as the Infusoria by Wrisberg in 1765. In the mid-18th century, while Carl von Linnaeus largely ignored the protists, his contemporary Otto Friedrich Müller was the first to introduce protists to the binomial system of nomenclature. The Infusoria included not only protists but also bacteria and many groups of small invertebrate animals.
In the early 19th century, the German naturalist Georg August Goldfuss introduced the word “Protozoa” (early animals) as a class within Kingdom Animalia, to refer to three groups of very different organisms: Infusoria (the modern ciliates of today), Lithozoa (corals), Phytozoa (such as Cryptomonas) and Medusinae (jellyfish). Later, in 1845, Carl Theodor von Siebold was the first to establish Protozoa as a phylum of exclusively “unicellular animals” consisting of two classes: Infusoria (ciliates) and Rhizopoda (amoebae, foraminifera). Other scientists, such as Louis Agassiz, did not consider all of these organisms to be part of the animal kingdom, and by the middle of the century they were generally regarded within the groupings of Protozoa (early animals), Protophyta (early plants), Phytozoa (animal-like plants) and Bacteria (mostly considered plants). Microscopic organisms were increasingly constrained in the dichotomy between plant and animal. In 1858, the palaeontolgist Richard Owen was the first to define Protozoa as a separated kingdom of eukaryotic organisms, with “nucleated cells” and the “common organic characters” of plants and animals, although he also included sponges within this definition.
Origin of Kingdom Protista or Protoctista

In 1860, naturalist John Hogg proposed Protoctista (first-created beings) as the name for a fourth kingdom of nature, “Regnum Primigenum” (primigenal kingdom), the other kingdoms being Linnaeus' plant, animal and mineral. This kingdom comprised all the lower, primitive organisms, including Protophyta, Protozoa and Armophoctista (sponges), at the merging bases of the plant and animal kingdoms.
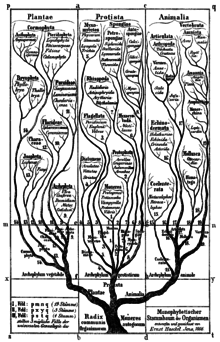
In 1866 the 'father of protistology', Ernst Haeckel, addressed the problem of classifying all these organisms as a mixture of animal and vegetable characters, and proposed Protistenreich (Kingdom Protista) as the third kingdom of life, comprising primitive forms that were “neither animals nor plants”. He grouped both prokaryotes and eukaryotes, both unicellular and multicellular organisms, as Protista. He retained the Infusoria in the animal kingdom, until Otto Butschli demonstrated that they were unicellular. At first, he included sponges and fungi, but in later publications he explicitly restricted Protista to predominantly unicellular organisms or colonies incapable of forming tissues. Within Protista he also retained bacteria, first as Mychota and then as Monera. He clearly separated Protista from true animals and plants on the basis that the defining character of protists was the absence of sexual reproduction, while the defining character of animals was the blastula stage of animal development. He also returned the terms protozoa and protophyta as subkingdoms of Protista.
Otto Butschli considered the kingdom to be too polyphyletic and rejected the inclusion of bacteria. He fragmented the kingdom into protozoa (only nucleated, unicellular animal-like organisms), while bacteria and the protophyta were a separate grouping. This strengthened the old concept of protozoa/protophyta from von Siebold, and the German naturalists asserted this view over the worldwide scientific community by the turn of the century. However, C. Clifford Dobell in 1911 brought attention to the fact that protists functioned very differently compared to the animal and vegetable cellular organization, and gave importance to Protista as a group with a different organization that he called “acellularity”, shifting away from the dogma of German cell theory. He coined the term Protistology and solidified it as a branch of study.
In 1938, Herbert Copeland resurrected Hogg's label, arguing that Haeckel's term Protista included anucleated microbes such as bacteria, which the term Protoctista (meaning "first established beings") did not. Under his four-kingdom classification (Monera, Protoctista, Plantae, Animalia), the protists and bacteria were finally split apart, recognizing the difference between anucleate (prokaryotic) and nucleate (eukaryotic) organisms. To firmly separate protists from plants and animals, he followed Haeckel's blastular definition of true animals, and proposed defining true plants as those with chlorophyll a and b, carotene, xanthophyll and production of starch. He also was the first to recognize that the unicellular/multicellular dichotomy was invalid. Still, he kept fungi within Protoctista, together with red algae, brown algae and protozoans. This classification was the basis for Whittaker's later definition of Fungi, Animalia, Plantae and Protista as the four kingdoms of life..
In the popular five-kingdom scheme published by Robert Whittaker in 1969, Protista was defined as eukaryotic “organisms which are unicellular or unicellular-colonial and which form no tissues”. Just as the prokaryotic/eukaryotic division was becoming mainstream, Whittaker, after a decade from Copeland's system, recognized the fundamental division of life between the prokaryotic Monera and the eukaryotic kingdoms: Animalia (ingestion), Plantae (photosynthesis), Fungi (absorption) and the remaining Protista.
In the five-kingdom system of Lynn Margulis, the term “protist” was reserved for microscopic organisms, while the more inclusive kingdom Protoctista (or protoctists) included certain large multicellular eukaryotes, such as kelp, red algae, and slime molds. Some use the term protist interchangeably with Margulis's protoctist, to encompass both single-celled and multicellular eukaryotes, including those that form specialized tissues but do not fit into any of the other traditional kingdoms.
Phylogenetics and the modern definition
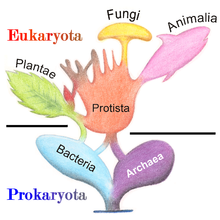
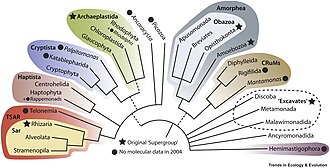
The five-kingdom models remained the accepted classification until the development of molecular phylogenetics in the late 20th century, when it became apparent that neither protists nor monera were single groups of related organisms (they were not monophyletic groups), and the three-domain system (Bacteria, Archaea, Eukarya) became prevalent.
Today, Protista is not treated as a formal taxon, but the term "protist" is still commonly used for convenience in two ways. The most popular contemporary definition is a phylogenetic one, that recognizes protists as a paraphyletic group: a protist is any eukaryote that is not an animal, (land) plant, or (true) fungus; this definition excludes many unicellular groups, like the Microsporidia, Chytridiomycetes and yeast (fungi), and a non-unicellular group included in Protista in the past, the Myxozoa (animals).
The other definition describes protists primarily by functional or biological criteria: protists are essentially those eukaryotes that are never multicellular, that either exist as independent cells, or if they occur in colonies, do not show differentiation into tissues.
Because the protists are paraphyletic, the monophyletic kingdoms Animalia, Plantae and Fungi evolved from them. The newest classification systems of eukaryotes do not recognize the formal taxonomic ranks (phylum, class, order...) and instead only recognize the group that are clades of related organisms. This is intended to make the classification more stable in the long term and easier to update. In this new cladistic scheme, the protists are divided into wide branches or supergroups, such as: the SAR supergroup (Stramenopiles + Alveolata + Rhizaria), Opisthokonta (animals, fungi and all related protists), Archaeplastida (true plants and related protists), Amoebozoa (containing slime molds), Discoba (containing most excavates), and others.
Protozoa and Chromista
There is, however, one classification of protists based on traditional ranks that lasted until the 21st century. The protozoologist Thomas Cavalier-Smith, since 1998, developed a six-kingdom model (Bacteria, Animalia, Plantae, Fungi and the two protist kingdoms Protozoa and Chromista). In his context, paraphyletic groups take preference over clades: the "Protozoa" consist of monophyletic phyla such as Amoebozoa, Euglenozoa and Percolozoa, but also paraphyletic phyla such as "Choanozoa" (the opisthokont protists) and "Eolouka". In the same manner, kingdom "Chromista" contains many monophyletic phyla of protists such as Cercozoa and Gyrista, as well as paraphyletic groups such as "Hacrobia". This scheme endured until 2021, with his last publication where he revised Protozoa.
Reproduction
Protists generally reproduce asexually under favorable environmental conditions, but tend to reproduce sexually under stressful conditions, such as starvation or heat shock. Oxidative stress, which leads to DNA damage, also appears to be an important factor in the induction of sex in protists.
The demonstration of sex in protists
Eukaryotes emerged in evolution more than 1.5 billion years ago. The earliest eukaryotes were protists. Although sexual reproduction is widespread among multicellular eukaryotes, it seemed unlikely until recently, that sex could be a primordial and fundamental characteristic of eukaryotes. The main reason for this view was that sex appeared to be lacking in certain pathogenic protists whose ancestors branched off early from the eukaryotic family tree. However, several of these "early-branching" protists that were thought to predate the emergence of meiosis and sex (such as Giardia lamblia and Trichomonas vaginalis) are now known to descend from ancestors capable of meiosis and meiotic recombination, because they have a set core of meiotic genes that are present in sexual eukaryotes. Most of these meiotic genes were likely present in the common ancestor of all eukaryotes, which was likely capable of facultative (non-obligate) sexual reproduction.
This view was further supported by a 2011 study on amoebae. Amoebae have been regarded as asexual organisms, but the study describes evidence that most amoeboid lineages are ancestrally sexual, and that the majority of asexual groups likely arose recently and independently. Even in the early 20th century, some researchers interpreted phenomena related to chromidia (chromatin granules free in the cytoplasm) in amoebae as sexual reproduction.
Sexual reproduction in pathogenic protists
Some commonly found protist pathogens such as Toxoplasma gondii are capable of infecting and undergoing asexual reproduction in a wide variety of animals – which act as secondary or intermediate host – but can undergo sexual reproduction only in the primary or definitive host (for example: felids such as domestic cats in this case).
Some species, for example Plasmodium falciparum, have extremely complex life cycles that involve multiple forms of the organism, some of which reproduce sexually and others asexually. However, it is unclear how frequently sexual reproduction causes genetic exchange between different strains of Plasmodium in nature and most populations of parasitic protists may be clonal lines that rarely exchange genes with other members of their species.
The pathogenic parasitic protists of the genus Leishmania have been shown to be capable of a sexual cycle in the invertebrate vector, likened to the meiosis undertaken in the trypanosomes.
Ecology

Free-living protists occupy almost any environment that contains liquid water. Many protists, such as algae, are photosynthetic and are vital primary producers in ecosystems, particularly in the ocean as part of the plankton. Other protists include pathogenic species, such as the kinetoplastid Trypanosoma brucei, which causes sleeping sickness, and species of the apicomplexan Plasmodium, which cause malaria.
Biomass
Protists make up a large portion of the biomass in both marine and terrestrial ecosystems. It has been estimated that protists account for 4 gigatons (Gt) of biomass in the entire planet Earth. This amount is smaller than 0.01% of all biomass, but is still double the amount estimated for all animals (2 Gt). Together, protists, animals, archaea (7 Gt) and fungi (12 Gt) only account for less than 10% of the total biomass of the planet, because plants (450 Gt) and bacteria (70 Gt) are the remaining 80% and 15% respectively.
Metabolism
Nutrition can vary according to the type of protist. Most eukaryotic algae are autotrophic, but the pigments were lost in some groups. Other protists are heterotrophic, and may present phagotrophy, osmotrophy, saprotrophy or parasitism. Some are mixotrophic. Some protists that do not have / lost chloroplasts/mitochondria have entered into endosymbiontic relationship with other bacteria/algae to replace the missing functionality. For example, Paramecium bursaria and Paulinella have captured a green alga (Zoochlorella) and a cyanobacterium respectively that act as replacements for chloroplast. Meanwhile, a protist, Mixotricha paradoxa that has lost its mitochondria uses endosymbiontic bacteria as mitochondria and ectosymbiontic hair-like bacteria (Treponema spirochetes) for locomotion.
Many protists are flagellate, for example, and filter feeding can take place where flagellates find prey. Other protists can engulf bacteria and other food particles, by extending their cell membrane around them to form a food vacuole and digesting them internally in a process termed phagocytosis.
| Nutritional type | Source of energy | Source of carbon | Examples |
|---|---|---|---|
| Photoautotrophs | Sunlight | Organic compounds or carbon fixation | Most algae |
| Chemoheterotrophs | Organic compounds | Organic compounds | Apicomplexa, Trypanosomes or Amoebae |
For most important cellular structures and functions of animal and plants, it can be found a heritage among protists.
Parasitism: role as pathogens
Some protists are significant parasites of animals (e.g.; five species of the parasitic genus Plasmodium cause malaria in humans and many others cause similar diseases in other vertebrates), plants (the oomycete Phytophthora infestans causes late blight in potatoes) or even of other protists. Protist pathogens share many metabolic pathways with their eukaryotic hosts. This makes therapeutic target development extremely difficult – a drug that harms a protist parasite is also likely to harm its animal/plant host. A more thorough understanding of protist biology may allow these diseases to be treated more efficiently. For example, the apicoplast (a nonphotosynthetic chloroplast but essential to carry out important functions other than photosynthesis) present in apicomplexans provides an attractive target for treating diseases caused by dangerous pathogens such as plasmodium.
Recent papers have proposed the use of viruses to treat infections caused by protozoa.
Researchers from the Agricultural Research Service are taking advantage of protists as pathogens to control red imported fire ant (Solenopsis invicta) populations in Argentina. Spore-producing protists such as Kneallhazia solenopsae (recognized as a sister clade or the closest relative to the fungus kingdom now) can reduce red fire ant populations by 53–100%. Researchers have also been able to infect phorid fly parasitoids of the ant with the protist without harming the flies. This turns the flies into a vector that can spread the pathogenic protist between red fire ant colonies.
Fossil record
Many protists have neither hard parts nor resistant spores, and their fossils are extremely rare or unknown. Examples of such groups include the apicomplexans, most ciliates, some green algae (the Klebsormidiales),choanoflagellates,oomycetes,brown algae,yellow-green algae,Excavata (e.g., euglenids). Some of these have been found preserved in amber (fossilized tree resin) or under unusual conditions (e.g., Paleoleishmania, a kinetoplastid).
Others are relatively common in the fossil record, as the diatoms,golden algae,haptophytes (coccoliths),silicoflagellates, tintinnids (ciliates), dinoflagellates,green algae,red algae,heliozoans, radiolarians,foraminiferans,ebriids and testate amoebae (euglyphids, arcellaceans). Some are even used as paleoecological indicators to reconstruct ancient environments.
More probable eukaryote fossils begin to appear at about 1.8 billion years ago, the acritarchs, spherical fossils of likely algal protists. Another possible representative of early fossil eukaryotes are the Gabonionta.
See also
Bibliography
General
- Haeckel, E. Das Protistenreich. Leipzig, 1878.
- Hausmann, K., N. Hulsmann, R. Radek. Protistology. Schweizerbart'sche Verlagsbuchshandlung, Stuttgart, 2003.
- Margulis, L., J.O. Corliss, M. Melkonian, D.J. Chapman. Handbook of Protoctista. Jones and Bartlett Publishers, Boston, 1990.
- Margulis, L., K.V. Schwartz. Five Kingdoms: An Illustrated Guide to the Phyla of Life on Earth, 3rd ed. New York: W.H. Freeman, 1998.
- Margulis, L., L. Olendzenski, H.I. McKhann. Illustrated Glossary of the Protoctista, 1993.
- Margulis, L., M.J. Chapman. Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth. Amsterdam: Academic Press/Elsevier, 2009.
- Schaechter, M. Eukaryotic microbes. Amsterdam, Academic Press, 2012.
Physiology, ecology and paleontology
- Foissner, W.; D.L. Hawksworth. Protist Diversity and Geographical Distribution. Dordrecht: Springer, 2009
- Fontaneto, D. Biogeography of Microscopic Organisms. Is Everything Small Everywhere? Cambridge University Press, Cambridge, 2011.
- Levandowsky, M. Physiological Adaptations of Protists. In: Cell physiology sourcebook : essentials of membrane biophysics. Amsterdam; Boston: Elsevier/AP, 2012.
- Moore, R. C., and other editors. Treatise on Invertebrate Paleontology. Protista, part B (vol. 1, Charophyta, vol. 2, Chrysomonadida, Coccolithophorida, Charophyta, Diatomacea & Pyrrhophyta), part C (Sarcodina, Chiefly "Thecamoebians" and Foraminiferida) and part D (Chiefly Radiolaria and Tintinnina). Boulder, Colorado: Geological Society of America; & Lawrence, Kansas: University of Kansas Press.
External links
- A java applet for exploring the new higher level classification of eukaryotes
- Holt, Jack R. and Carlos A. Iudica. (2013). Diversity of Life. http://comenius.susqu.edu/biol/202/Taxa.htm. Last modified: 11/18/13.
- Plankton Chronicles – Protists – Cells in the Sea – video
- Tree of Life: Eukaryotes
- Tsukii, Y. (1996). Protist Information Server (database of protist images). Laboratory of Biology, Hosei University.[1]. Updated: March 22, 2016.